研究
一開始就可以使用突觸
我們的大腦是視覺信息、聽力信息、練習每個功能,例如運動功能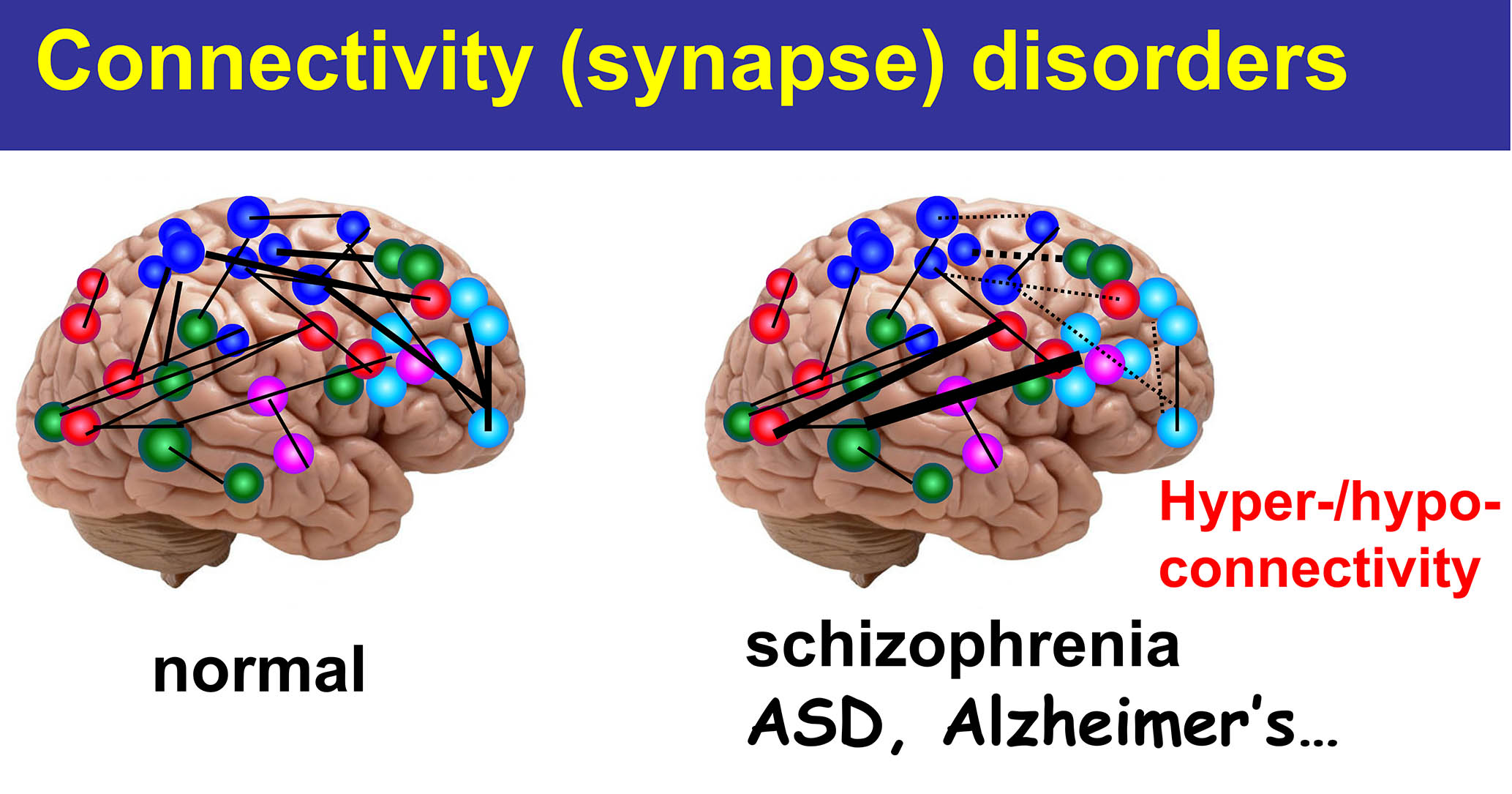 它由神經網絡組成。。神經網絡是、大約1000億個神經元、它們是通過通過一個稱為“突觸”的結相互連接而形成的。。每個神經元平均有大約10,000個突觸。、突觸的數量估計達到1,000萬億。這些突觸如何形成?最新研究結果的結果、突觸不僅是根據遺傳信息形成的、通過環境因素,出生後的經驗和學習,在整個生命中繼續在功能和形態上繼續變化。突觸是、這是使我們人類的根源。實際的、根據最近的研究結果、抑鬱症和精神分裂症等精神疾病、自閉症譜系障礙等發育障礙、此外,包括阿爾茨海默氏病在內的癡呆等神經疾病被認為是“突觸疾病”。。
它由神經網絡組成。。神經網絡是、大約1000億個神經元、它們是通過通過一個稱為“突觸”的結相互連接而形成的。。每個神經元平均有大約10,000個突觸。、突觸的數量估計達到1,000萬億。這些突觸如何形成?最新研究結果的結果、突觸不僅是根據遺傳信息形成的、通過環境因素,出生後的經驗和學習,在整個生命中繼續在功能和形態上繼續變化。突觸是、這是使我們人類的根源。實際的、根據最近的研究結果、抑鬱症和精神分裂症等精神疾病、自閉症譜系障礙等發育障礙、此外,包括阿爾茨海默氏病在內的癡呆等神經疾病被認為是“突觸疾病”。。
我們正在研究什麼?
取決於神經活動、突觸時信號傳遞效率的長期提高(長期增強):長期增強 (LTP))和衰落現象(長期抑制):長期抑鬱症 (有限公司))已知。這種變化是、它被認為是短期至中期記憶的實體(在人類中,每天的單位)。。另一方面、較長的記憶(一個月)、據信,突觸本身是由形態學變化(例如新形成或消失)進行的。在Yuzaki研究所、這兩個現象:1)突觸的功能變化(LTP/LTD)、②形態變化,例如突觸形成,維護和去除、什麼分子機制是、我們旨在了解它們如何參與個人層面上神經迴路和行為的變化。。
這到底有什麼幫助?
突觸如何在功能上改變、或如何形成突觸、維護它、會丟失嗎?、澄清過程、了解由突觸異常引起的各種心理和神經系統疾病以及發育障礙的病理。、這對於開發新治療非常重要。各種發展是可能的,例如:。但、什麼很重要、更好地了解突觸處發生的現象這是、我相信該申請將在此之後遵循。
- 我們通過闡明為什麼只能在幼兒(甚至三胞胎的第100個靈魂)才能實現某些學習來培養健康的大腦。
- 通過闡明引起精神疾病和發育障礙的突觸的異常、連接到新的基本診斷和治療方法
- 在神經元死亡發生之前、通過在突觸水平上進行干預來防止癡呆症的進展
- 通過移植IPS細胞衍生的神經元的再生醫學。、通過促進突觸形成增加恢復。
您專門從事什麼樣的研究?…
1)使用光遺傳學
研究LTP/LTD機制
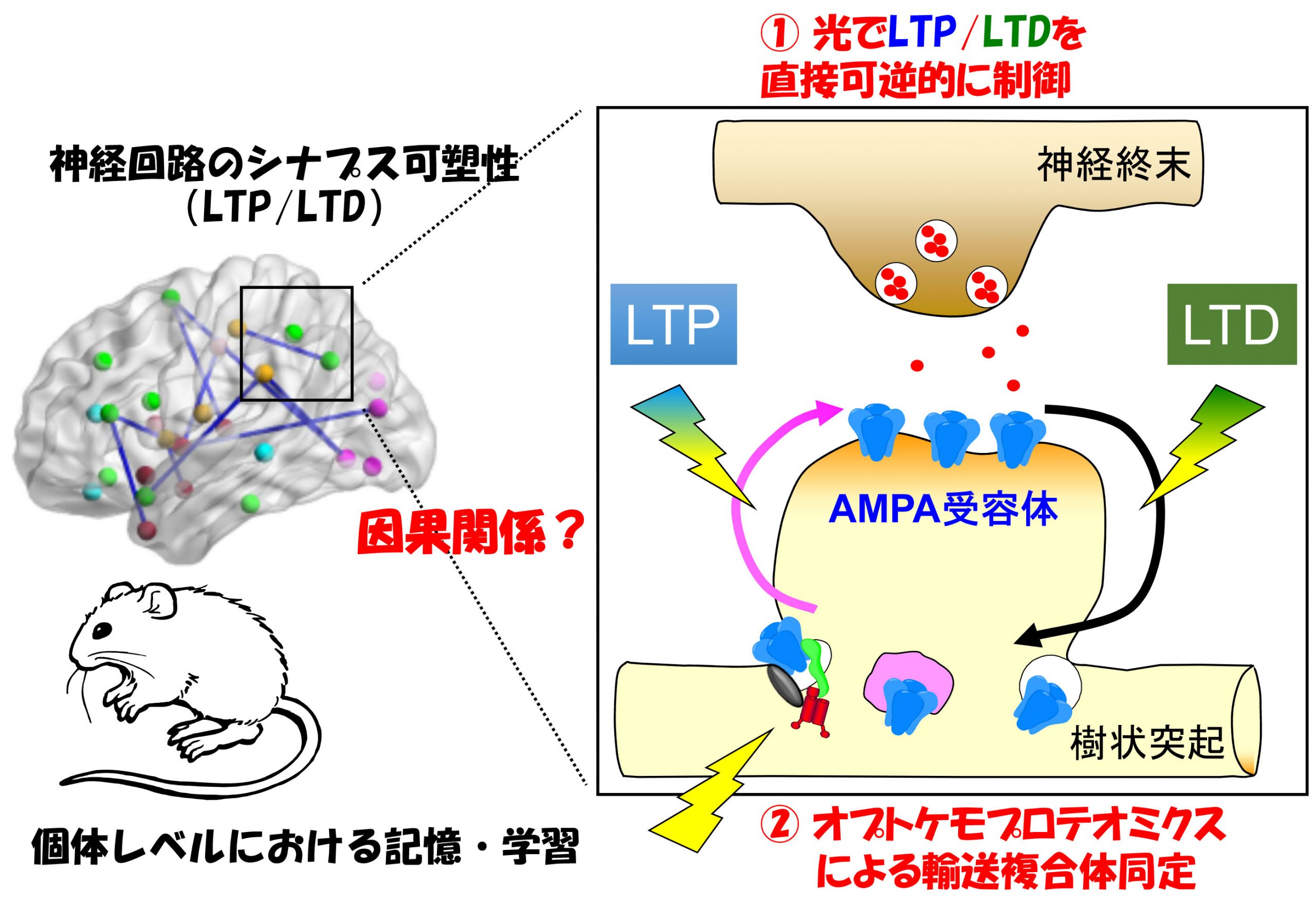
響應神經活動的突觸可塑性現象、長期增強是響應於人為地應用於突觸的電刺激的響應(LTP)和長期抑制(有限公司)使用該模型對其進行了多年的研究。但、以這種方式造成的LTP/LTD是、在個人層面上,記憶與學習之間確實存在因果關係嗎?、這個基本問題仍未解決。我們、通過光刺激、通過開發一種可以直接在目標突觸下敏銳而可逆的LTD的光遺傳學工具(Photonsaber)、我們成功地表明了小腦中的運動學習是由有限公司(Neuron)進行的, 2018()。通過進一步開發此類工具,、我們旨在闡明在短期和中期記憶中發揮作用的分子機制。。
2)研究以補體家族為中心的突觸形成分子
補體C1q是一種識別先天免疫系統中異物的分子,是一種進化,非常古老的分子。。有趣的東西、類似於C1q(補體家族分子)的一組分子不僅是免疫系統、我們發現它調節中樞神經系統中的突觸形成和成熟過程。
在小腦中、平行纖維,顆粒細胞的軸突,在Purkinje細胞中形成樹突和突觸。平行纖維分泌補體家族分子CBLN1。、同時,它與平行纖維中存在的神經毒素(NRX)結合、purkinje細胞中存在的Δ2谷氨酸受體型(glud2)它也與。這個由NRX-CBLN1-GLUD2組成的三方復合物、我們報告說,平行纖維和浦肯野細胞之間的突觸粘附至關重要(科學 2010; 2016()。攀岩纖維,下橄欖核的軸突,也形成了浦肯野細胞中的樹突和突觸。。C1QL1是一種補體的家族分子,是從攀岩纖維中分泌的。、我們發現,通過與Purkinje細胞中存在的七跨膜粘附分子BAI3結合,我們調節了鏟子纖維纖維 - - 純荷基細胞突觸的成熟和維持(Neuron 2014)。。平行纖維-Purkinje細胞、在兩種類型的突觸中,Tosho纖維Purkinje細胞、CBLN1, 釋放了不同補體的家族分子C1QL1。、競爭在Purkinje細胞的樹突上引起突觸形成是非常有趣的。。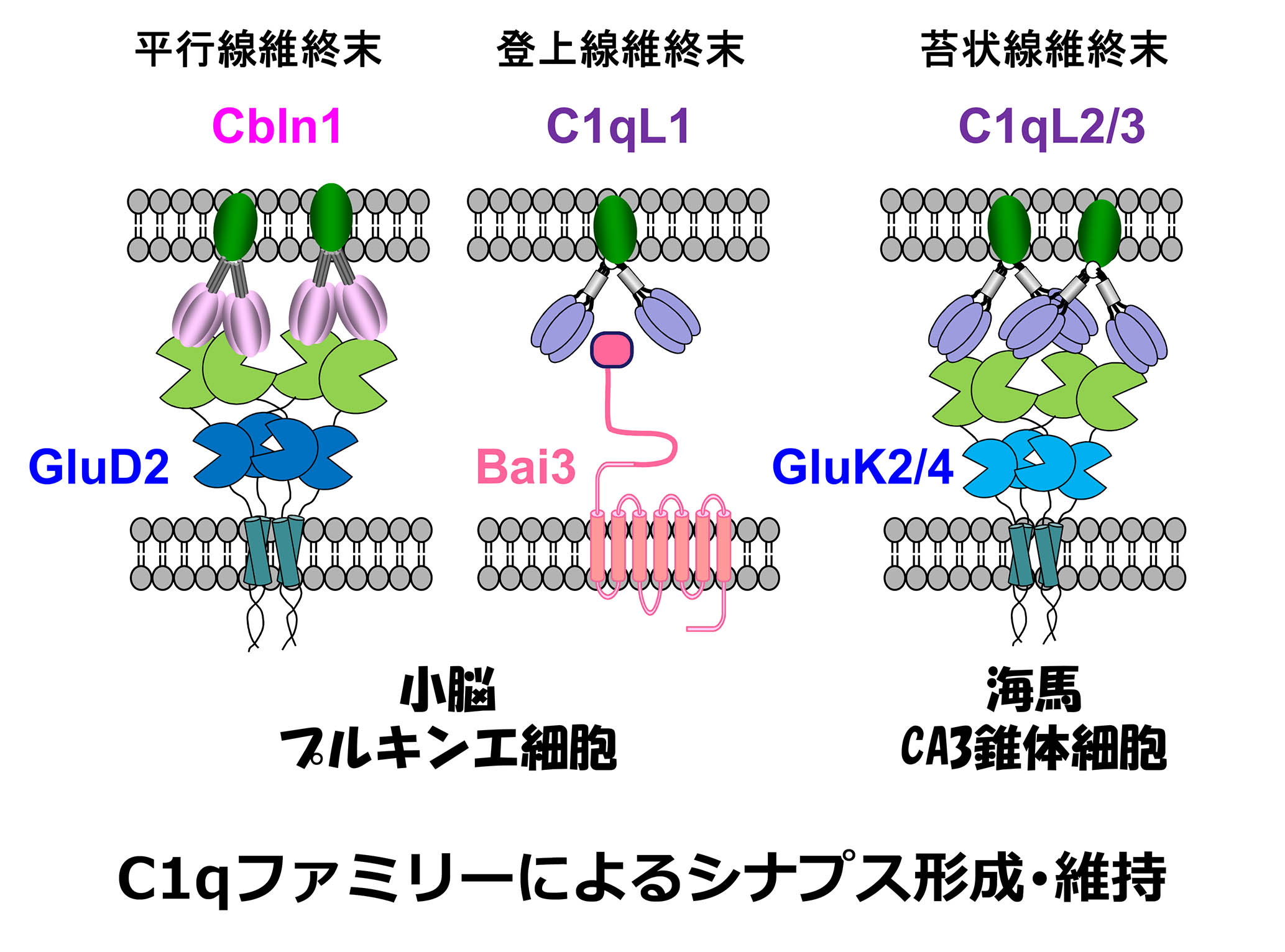
谷氨酸受體負責我們大腦中的興奮性神經傳遞。、在谷氨酸受體中,AMPA受體提供快速的神經傳遞。、海藻酸受體負責緩慢的傳播。海藻受體在海馬CA3錐體細胞中高度表達。、特別是通過在苔蘚纖維(齒狀回顆粒細胞的軸突)產生的後突觸中積聚。、它在引起時間和空間添加神經活動方面起著重要作用。另一方面、在顳葉癲癇中,由於其成癮,這種海藻酸受體在癲癇病局灶性形成中起著重要作用。
我會做的。到目前為止,這是一個完全的謎,為什麼海藻酸受體在後苔蘚纖維突觸中積聚。。我們是從苔蘚纖維分泌的補體C1QL2和C1QL3的補體分子。、我們發現,這是一個分子,它決定了在CA3錐體細胞(神經元)中突觸後突觸中的海藻酸受體的積累(神經元), 2016()。補體家族分子是由神經活動合成的。、有分泌的財產、因為它與炎症密切相關、預計將參與各種疾病的病理。
3)開發人造突觸連接器
構成大腦各種神經迴路的突觸、通過闡明形成什麼突觸分子並維持什麼、您可以開發具有自然界不存在的屬性的人造突觸連接器。迄今為止、全部
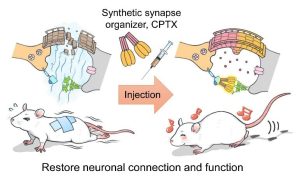
我們已經闡明了與顱神經迴路中存在於興奮性突觸中存在的谷氨酸受體1(NPTX1)的晶體結構。、通過將其結構與CBLN1相結合,CBLN1在小腦中具有很強的突觸能力、新的人造突觸組織者CPTX(=cBLN1 + nptx1)成功開發(科學2020)。CPTX是
即使在小腦以外的其他神經迴路中,它也具有強烈連接突觸的能力。、CPTX共濟失調、阿爾茨海默氏病、當給脊髓損傷的模型小鼠施用時、突觸將在幾天內形成、改進的協調、恢復學習和記憶功能、恢復癱瘓的後腿的運動功能,等等。、我們能夠顯著改善每個條件。。通過開發這種人造突觸連接器、自閉症譜系障礙、精神分裂症、預計將導致為突觸疾病(例如阿爾茨海默氏病)創建和應用新的治療策略。
—英語版本—
一開始是突觸
大約 100 十億個神經元通過“突觸”相互連接,以形成人腦的各種神經迴路. 雖然突觸的數量可以達到 1,000 百萬, 如何在電路中實際形成突觸- 和神經元特異性的方式仍然不完全理解. 最近的成像研究表明,突觸不僅是由遺傳信息形成的, 但也通過一生的神經元活動不斷修改. 此外, 越來越清楚的是許多神經發育, 神經精神病和神經系統疾病, 例如自閉症譜系障礙, 精神分裂症和阿爾茨海默氏病, 是由突觸功能缺陷引起的. 所以, 必須闡明如何形成突觸是至關重要的, 維護和修改以更好地了解我們的大腦在生理和病理條件下如何功能和功能障礙, 分別. 朝著這個目標, 我們關注的兩個主要項目如下:
我們研究什麼?
1) 神經元活動功能突觸連通性的變化
突觸連通性通過神經元活動的變化在功能上修改, 一種稱為突觸可塑性的現象. 長期增強 (LTP) 和長期抑鬱症 (有限公司) 興奮性神經傳遞已被深入研究,作為介導學習和記憶的突觸可塑性模型. 雖然已經很明顯LTP和LTD主要是由於活動引起的突觸後AMPA型谷氨酸受體的增加和減少引起的 (ampars) 直接介導快速興奮性神經傳遞, 分別, 尚不完全清楚地與腦功能有因果關係,與大腦功能有關,介導了體內的特定行為變化. 解決這個問題, Yuzaki實驗室正在開發光遺傳學工具,該工具可以直接調節活物中的LTP和LTD.
2) 突觸組織者的結構突觸連通性的變化
調節突觸形成的分子, 維護和修改稱為突觸組織者. 他們分為兩個主要課程, 細胞粘附分子, 例如神經蛋白和神經素, 並分泌的擴散因素, 例如Wnt和FGF. 有趣的是, 某些分泌的因素保留在突觸裂縫處的細胞外基質中,並作為pre的細胞外支架- 和突觸後蛋白, 因此被稱為細胞外支架蛋白 (ESPS). 突觸組織者的ESP型包括神經元五肽和C1Q家族蛋白, 其特徵是存在C末端球狀C1Q域. C1Q, C1Q家族的原型, 屬於補充的先天免疫系統,並認識到各種異物. Yuzaki實驗室發現C1Q家族蛋白, 例如小腦 (cblns) 和C1Q樣蛋白 (C1QLS) 主要在中樞神經系統中表達,並且一直在表徵其在突觸時的功能.
—
最近的代表論文 (自從 2005 ):
有關詳細信息:  https://orcid.org/0000-0002-5750-3544
https://orcid.org/0000-0002-5750-3544
- Kingawa W *, Paterini關閉, 松本K。, 伊莎貝爾AM, iida i, miura e, Nozawa K, Yamasaki t, Sakimura K, Yuzaki M **, 長的. 海藻酸鹽受體通過與小腦的突觸組織者形成複合物來調節突觸完整性和可塑性. 細胞報告 43:114427, 2024.
- Itoh m, Piot l, Monly L, Paolotti P *, Yuzaki M.*缺乏GLUD受體直接配體門控離子通道活性的證據 美國科學院校 121:E2406655121, 2024.
- Nozawa K, sogabe t, Hayashi a, Motohashi J, miura e, Arai i, Yuzaki M*. 神經膠質配體的體內納米那鏡景觀,依次突觸規範. 神經元 110:3168-3185, 2022..
- Ojima K†, 金卡瓦W†, Yamasaki t, miura和, Itoh m, Michibata y Michibata和, Kubota r, 杜拉, miura e, Naona h, 美津, 高橋, Yuzaki M*, hamachi i *, kiyonaka s **. 用於激活腦組織中GPCR型谷氨酸受體的協調化學遺傳學. 常見的nat. 13(1):3167, 2022.
- 鈴木K†, Elegheert J†, 歌曲I†, Sasakura H†, SenkovO。, 松本K。, murning w, 克萊頓AJ, Chang Vt, Ferrer-Ferrer m, miura e, Kaushik r,Ikeno M, 莫里亞, Takeuchi y, Shimada t, 額頭s, Stoyanov S., 渡邊m, Takeuchi k, dityatev a*, Aricescu ar*, Yuzaki M ** . 合成突觸組織者蛋白恢復谷氨酸能神經元電路. 科學 369:EABB4853, 2020. (†聯合第一, *共同對應, **主要作者)
- 傳播k, kono m, Narumi S., Motohashi J, murning w, kohda k, Yuzaki m. 來自顆粒細胞軸突中溶酶體的突觸組織者CBLN1的活性依賴性分泌. 神經元 102:1184-1198, 2019.
- kono m, murning w, 吉田, Yuzaki m. NMDA受體中的神經元中的長期抑鬱和運動學習. J生理學. 597:903-920, 2019.
- murning w, kahoh a, Narumi S., miura e, Motohashi J, 高橋a, kohda k, 福川Y, Yuzaki M **, 松本S*。 突觸AMPA受體內吞作用的光遺傳學控制揭示了LTD在運動學習中的作用。神經元 99:985-998, 2018. (*共同對應的作者; **主要作者).
- Yuzaki m. 中樞神經系統中的兩類分泌的突觸組織者. Annu Rev Physiol80:243-262, 2018.
- 薩克t, Yamashita k, Hayashi k, Miyatake s, Tamura-Nakano m, 兩個h, Furuta和, Shioi g, miura e, takeo yh, 吉田, Yahikozawa h, 松本n, Yuzaki m, 鈴木. MTCL1在維持Purkinje神經元軸突初始段中起著至關重要的作用. Embo j 36:1227-1242, 2017.
- Wakayama s, Kiyonaka s, Arai i, murning w, 松本, 傳播k, Nemoto YL, Kusumi a, Yuzaki m, 哈馬奇. 用於可視化活神經元中天然AMPA受體的化學標記. 共同的性質 8:14850, 2017.
- Yuzaki m, Aricescu ar. 一個充滿成年的故事. 趨勢神經科學 40:138-150, 2017.
- 額頭s, Konno K., 安倍, Motohashi J, kohda k, Sakimura K, 渡邊m, Yuzaki m. CBLN1在小鼠非運動功能中的作用. J Neurosci, 36:11801-11816, 2016.
- Elegheert J, murning w, 粘土是, 小腿n, 又來了, 松本K。, kohda k, miura e, 羅斯曼, Mindakidis n, Motohashi J, Chang Vt, Siebold c, Greger IH, 中川T, Yuzaki M*, Aricescu ar*. 集成GLUD受體在突觸組織器中的結構基礎. 科學 353:295-299, 2016. (*共同對應作者).
- Kiyonaka s, Kubota r, Michibata y Michibata和, Sakakura m, 高橋h, 命名為t, inoue r, Yuzaki m, 哈馬奇. 使用活細胞內的配位化學對膜結合的谷氨酸受體的變構激活. 自然化學 8 :958-967, 2016.
- 松本K。, budentoso t, Mindakidis n, Suy, miura e, murning w, Yamasaki m, Konno K., Uchigashima m, 安倍, 渡邊i, 卡諾, 渡邊m, Sakimura K, Aricescu ar, Yuzaki m. C1Q樣蛋白的海藻酸鹽受體功能的反式突觸調節. 神經元 90:752-767, 2016.
- takeo yh, murning w, miura e, Yuzaki m. RORα調節體內小腦Purkinje細胞中樹突發育的多個方面. J Neurosci 35:12518-12534, 2015.
- murning w, Mindakidis n, miura e, 安倍, 松本K。, takeo yh, kohda k, Motohashi J, 高橋a, 命中S., Muramatsu Si, 渡邊m, Sakimura K, Aricescu ar, Yuzaki m. 為了確定和維持小鼠小腦中的單打攀岩纖維,需要順行C1QL1信號傳導. 神經元 85:316-329, 2015.
- 松本, murning w, budentoso t, 野村, kohda k, Yuzaki m. Stargazin在長期抑鬱症中調節通過銜接蛋白複合物調節AMPA受體運輸. 共同的性質 4:2759, 2013.
- kohda k, murning w, 松本, Yamamoto t, Hirano H, Yuzaki m. D2谷氨酸受體大門長期抑鬱,通過協調兩個AMPA受體磷酸化位點之間的相互作用. 美國科學院校 110:E948-957, 2013.
- ito-ishida a, 宮崎T, miura e, 松本K。, 渡邊m, Yuzaki M*, Okabe S*. 突觸前釋放的CBLN1在小腦突觸形成過程中與GLUD2相互作用會誘導動態軸突結構變化。神經元 76:549-564, 2012. (*共同對應作者).
- Unoki t, 松本, murning w, 來自UT, kohda k, 鈴木A。, funakoshi y, 長谷川h, Yuzaki M*, 你好. NMDA受體介導的PIP5K激活產生PI(4,5)P2對於LTD期間的AMPA受體內吞作用至關重要。神經元 73: 135-148, 2012. (*共同對應作者)
- murning w, Miyoshi y, 哈馬斯, k, 松本, 松本K。, kohda k, 我, Motohashi J, Konno r, Zaitsu k, Yuzaki M. D-絲氨酸通過Delta2谷氨酸受體調節小腦有限公司和運動配位。自然神經科學 14: 603-611, 2011.
- Yuzaki M. Cbln1及其家族蛋白在突觸形成和維護中。Curr Opin Neurobiol21:215-220, 2011.
- Jitsuki s, tamemoto, 川崎T, 塔達h, 高橋a, Becamel c, 健康, Yuzaki m, Zukin RS, Ziff EB, 凱塞爾(Kessels)HW, 高橋T. 5-羥色胺介導皮質迴路的跨模式重組。神經元 69: 780-92, 2011.
- 松本K。, miura e, Miyzaki t, murning w, 我, Narumi S., 福川Y, ito-ishida a, Kondo t, shigemoto r, 渡邊m, Yuzaki m. CBLN1是孤兒谷氨酸受體Δ2的配體, 雙向突觸組織者。科學 328: 363-368, 2010.
- Nishiyama J, 松本K。, murning w, Yamada n, Motohashi J, mizushima n, Yuzaki m. 在Lurcher小鼠中對神經退行性的重新評估: 組成型離子通量導致細胞死亡, 不是, 自噬。J Neurosci 30:2177-87, 2010.
- kakegawa, W., 宮崎駿, T., 點, K., 馬蘇達, K., EMI, K., Motohashi, J., 渡邊, M., 淺, m. Glud2的N末端結構域 (glurΔ2) 招募了突觸前末端並調節體內小腦突觸發生, J Neurosci 29:5738-5748, 2009.
- iijima, T., EMI。, K., 淺, m. CBLN1表達的活動依賴性抑制: 小腦突觸的發育和穩態調節機制。j. Neurosci29:5425-5434、2009.
- 松本, miura e, 松本K。, murning w, kohda k, 渡邊m, Yuzaki m. 缺乏銜接蛋白AP-4的神經元軸突自噬小體中AMPA受體的積累。神經元 57: 1-16, 2008.
- ito-ishida, 一個。, Miura, E., EMI, K., 馬蘇達, K., iijima, T., Kondo, T., 點, k. 渡邊, M., 淺, m. CBLN1調節成熟小腦Purkinje細胞中興奮性突觸的快速形成和維持體外 和徒然. J Neurosci 28:5920-5930, 2008.
- kakegawa, W., 宮崎駿, T., EMI, K., 馬蘇達, K., 點, K., Motohashi, J., 機器, M., 卡瓦哈拉, s, 渡邊, M., 淺, m. Glurdelta2的C末端PDZ結合基序對突觸可塑性和小腦運動學習的差異調節。J Neurosci 28:1460-1468, 2008.
- kakegawa,W., 點, K., 淺, m. Delta2“離子型”谷氨酸受體充當非離子受體,可控制小腦突觸可塑性。J生理學 584:89-96, 2007.
- kakegawa, W., 宮崎駿, T., hirai, H., Motohashi, J., 機器, M., 渡邊, M., 淺, m. 通道孔的Ca2+通透性對於Delta2谷氨酸受體調節突觸可塑性和運動配位並不是必需的。J生理學 579.3:729-735, 2007.
- hirai h, pang z, bao d, 宮崎T, 那, miura e, 帕里斯J。, y, 渡邊w, Yuzaki M*, 摩根吉. CBLN1對於小腦的突觸完整性和可塑性至關重要。自然神經科學 8: 1534-1541, 2005. (*共同對應作者)