研究
一开始就可以使用突触
我们的大脑是视觉信息、听力信息、练习每个功能,例如运动功能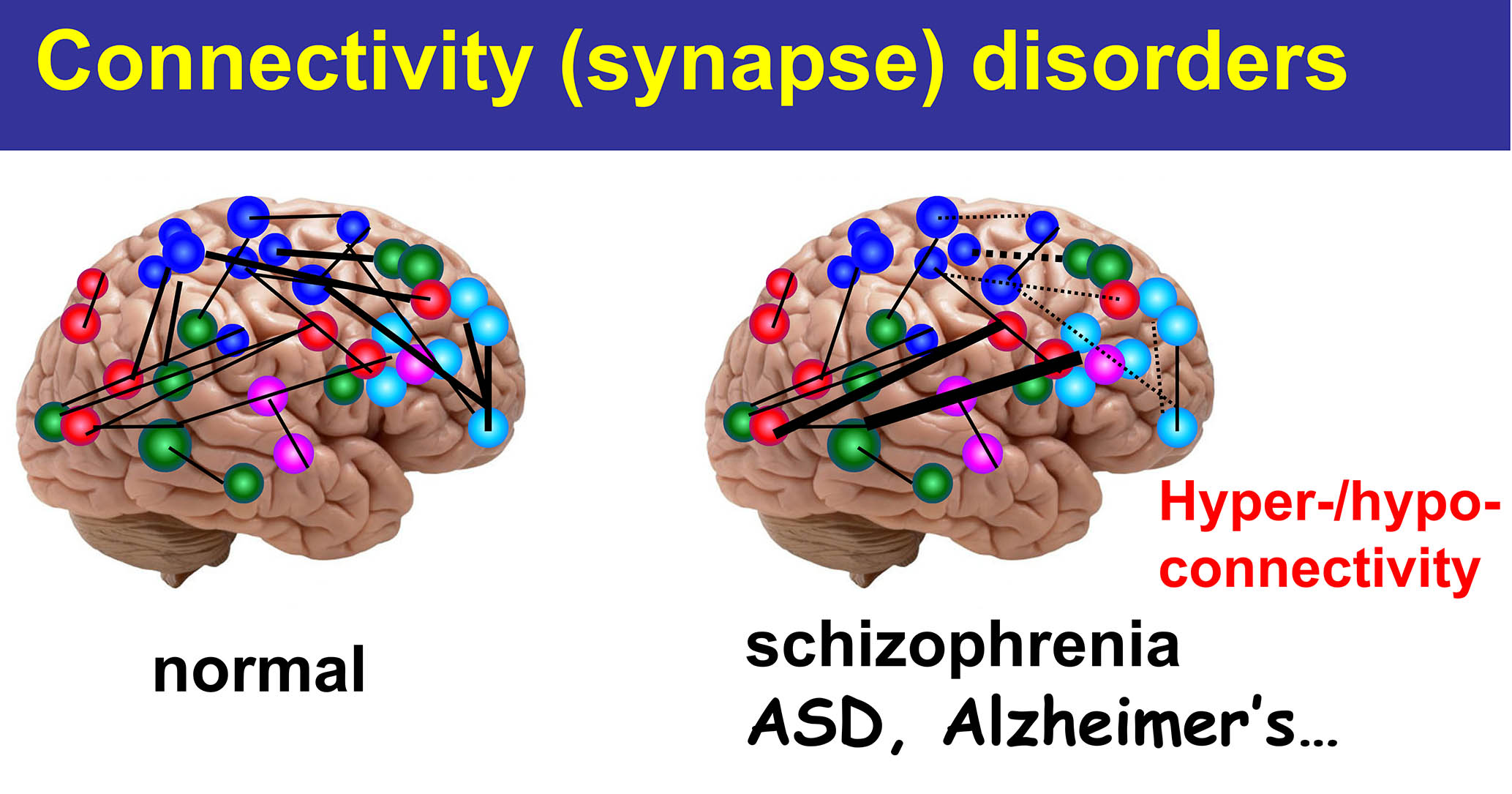 它由神经网络组成。。神经网络是、大约1000亿个神经元、它们是通过通过一个称为“突触”的结相互连接而形成的。。每个神经元平均有大约10,000个突触。、突触的数量估计达到1000万亿。这些突触如何形成?最新研究结果的结果、突触不仅是根据遗传信息形成的、通过环境因素,出生后的经验和学习,在整个生命中继续在功能和形态上继续变化。突触是、这是使我们人类的根源。实际的、根据最近的研究结果、抑郁症和精神分裂症等精神疾病、自闭症谱系障碍等发育障碍、此外,包括阿尔茨海默氏病在内的痴呆等神经疾病被认为是“突触疾病”。。
它由神经网络组成。。神经网络是、大约1000亿个神经元、它们是通过通过一个称为“突触”的结相互连接而形成的。。每个神经元平均有大约10,000个突触。、突触的数量估计达到1000万亿。这些突触如何形成?最新研究结果的结果、突触不仅是根据遗传信息形成的、通过环境因素,出生后的经验和学习,在整个生命中继续在功能和形态上继续变化。突触是、这是使我们人类的根源。实际的、根据最近的研究结果、抑郁症和精神分裂症等精神疾病、自闭症谱系障碍等发育障碍、此外,包括阿尔茨海默氏病在内的痴呆等神经疾病被认为是“突触疾病”。。
我们正在研究什么?
取决于神经活动、突触时信号传递效率的长期提高(长期增强):长期增强 (LTP))和衰落现象(长期抑制):长期抑郁症 (有限公司))已知。这种变化是、短期から中期記憶(ヒトの場合は時間~日の単位)の実体と考えられています。另一方面、较长的记忆(一个月)、据信,突触本身是由形态学变化(例如新形成或消失)进行的。在Yuzaki研究所、这两个现象:1)突触的功能变化(LTP/LTD)、②形态变化,例如突触形成,维护和去除、什么分子机制是、我们旨在了解它们如何参与个人层面上神经回路和行为的变化。。
这到底有什么帮助?
突触如何在功能上改变、或如何形成突触、维护它、会丢失吗?、澄清过程、了解由突触异常引起的各种心理和神经系统疾病以及发育障碍的病理。、这对于开发新治疗非常重要。各种发展是可能的,例如:。但、什么很重要、更好地了解突触处发生的现象这是、我相信该申请将在此之后遵循。
- 我们通过阐明为什么只能在幼儿(甚至三胞胎的100个三胞胎)才能在分子水平上实现某些学习来培养健康的大脑。
- 通过阐明引起精神疾病和发育障碍的突触的异常、连接到新的基本诊断和治疗方法
- 在神经元死亡发生之前、通过在突触水平上进行干预来防止痴呆症的进展
- 通过移植IPS细胞衍生的神经元的再生医学。、通过促进突触形成增加恢复。
您专门从事什么样的研究?…
1)使用光遗传学
研究LTP/LTD机制
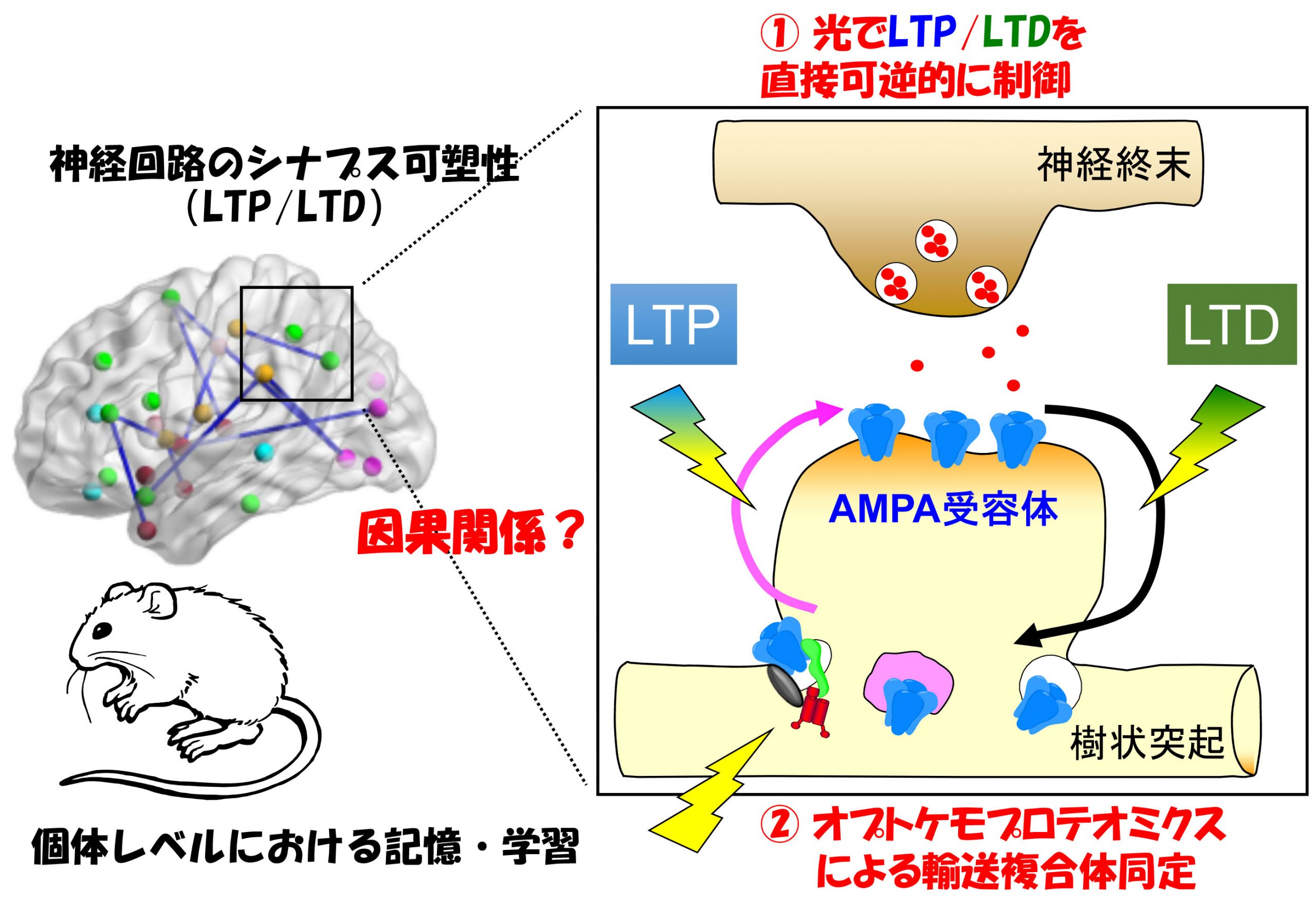
响应神经活动的突触可塑性现象、长期增强是响应于人为地应用于突触的电刺激的响应(LTP)和长期抑制(有限公司)使用该模型对其进行了多年的研究。但、以这种方式造成的LTP/LTD是、在个人层面上,记忆与学习之间确实存在因果关系吗?、这个基本问题仍未解决。我们、通过光刺激、通过开发一种可以直接在目标突触下敏锐而可逆的LTD的光遗传学工具(Photonsaber)、我们成功地表明了小脑中的运动学习是由有限公司(Neuron)进行的, 2018()。通过进一步开发此类工具,、我们旨在阐明在短期和中期记忆中发挥作用的分子机制。。
2)研究以补体家族为中心的突触形成分子
补体C1q是一种识别先天免疫系统中异物的分子,是一种进化,非常古老的分子。。有趣的东西、类似于C1q(补体家族分子)的一组分子不仅是免疫系统、我们发现它调节中枢神经系统中的突触形成和成熟过程。
在小脑中、平行纤维,颗粒细胞的轴突,在Purkinje细胞中形成树突和突触。平行纤维分泌补体家族分子CBLN1。、同时,它与平行纤维中存在的神经毒素(NRX)结合、purkinje细胞中存在的Δ2谷氨酸受体型(glud2)它也与。这个由NRX-CBLN1-GLUD2组成的三方复合物、我们报告说,平行纤维和浦肯野细胞之间的突触粘附至关重要(科学 2010; 2016()。攀岩纤维,下橄榄核的轴突,也形成了浦肯野细胞中的树突和突触。。C1QL1是一种补体的家族分子,是从攀岩纤维中分泌的。、我们发现,通过与Purkinje细胞中存在的七跨膜粘附分子BAI3结合,我们调节了铲子纤维纤维 - - 纯荷基细胞突触的成熟和维持(Neuron 2014)。。平行纤维-Purkinje细胞、在两种类型的突触中,Tosho纤维Purkinje细胞、CBLN1, 释放了不同补体的家族分子C1QL1。、竞争在Purkinje细胞的树突上引起突触形成是非常有趣的。。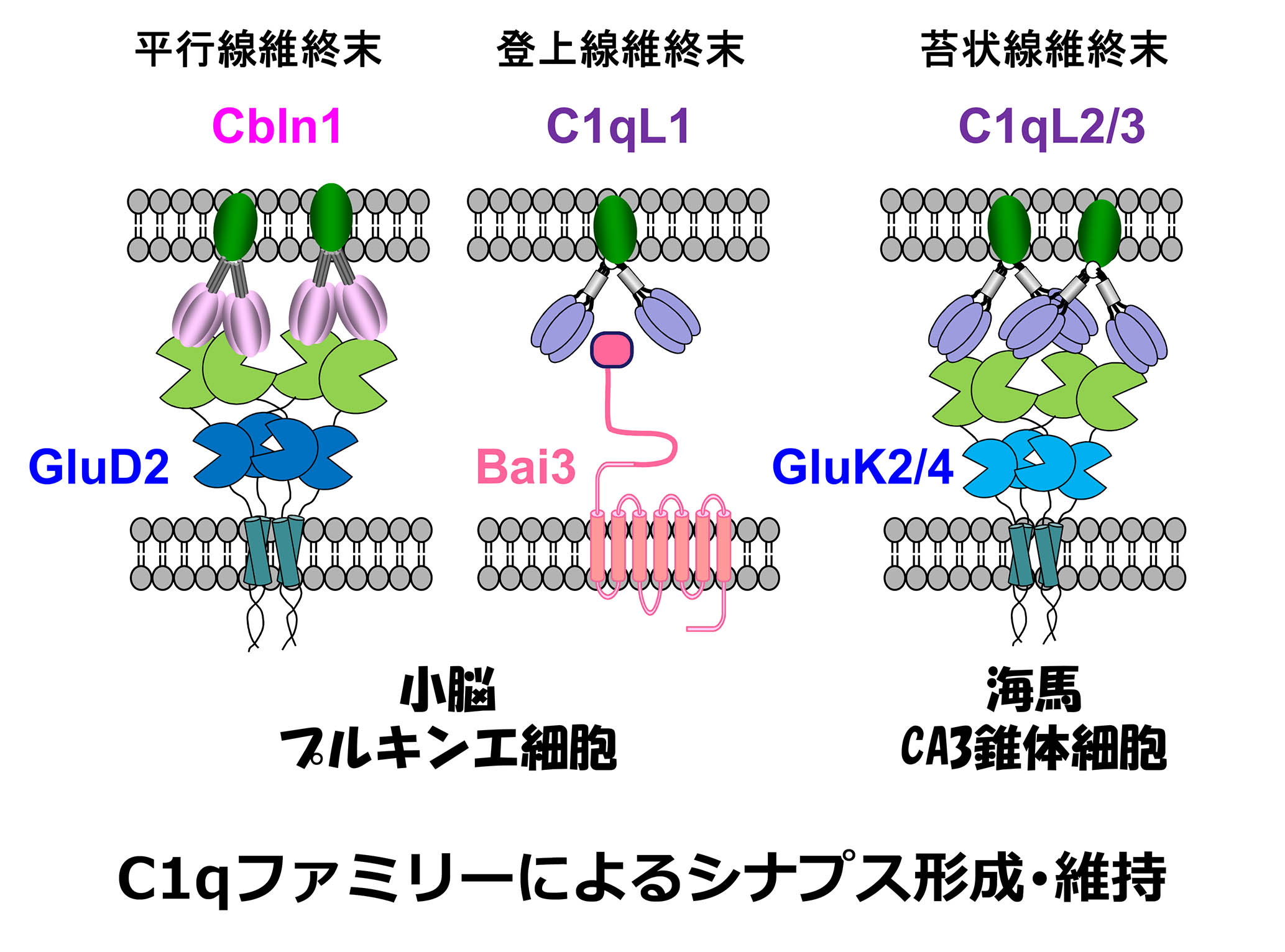
谷氨酸受体负责我们大脑中的兴奋性神经传递。、在谷氨酸受体中,AMPA受体提供快速的神经传递。、海藻酸受体负责缓慢的传播。海藻受体在海马CA3锥体细胞中高度表达。、特别是通过在苔藓纤维(齿状回颗粒细胞的轴突)产生的后突触中积聚。、它在引起时间和空间添加神经活动方面起着重要作用。另一方面、在颞叶癫痫中,由于其成瘾,这种海藻酸受体在癫痫病局灶性形成中起着重要作用。
我会做的。到目前为止,这是一个完全的谜,为什么海藻酸受体在后苔藓纤维突触中积聚。。我们是苔藓纤维分泌的补体家族分子C1QL2和C1QL3。、我们发现,这是一个分子,它决定了在CA3锥体细胞(神经元)中突触后突触中的海藻酸受体的积累(神经元), 2016()。补体家族分子是由神经活动合成的。、有分泌的财产、因为它与炎症密切相关、预计将参与各种疾病的病理。
3)开发人造突触连接器
构成大脑各种神经回路的突触、通过阐明形成和维护哪种突触形成分子、您可以开发具有自然界不存在的属性的人造突触连接器。迄今为止、全部
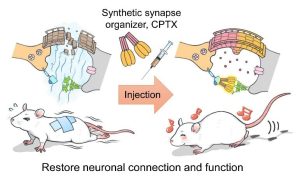
我们已经阐明了与颅神经回路中存在于兴奋性突触中存在的谷氨酸受体1(NPTX1)的晶体结构。、通过将其结构与CBLN1相结合,CBLN1在小脑中具有很强的突触能力、新的人造突触组织者CPTX(=cBLN1 + nptx1)成功开发(科学2020)。CPTX是
即使在小脑以外的其他神经回路中,它也具有强烈连接突触的能力。、CPTX共济失调、阿尔茨海默氏病、当给脊髓损伤的模型小鼠施用时、突触将在几天内形成、改进的协调、恢复学习和记忆功能、恢复瘫痪的后腿的运动功能,等等。、我们能够显着改善每个条件。。通过开发这种人造突触连接器、自闭症谱系障碍、精神分裂症、预计将导致为突触疾病(例如阿尔茨海默氏病)创建和应用新的治疗策略。
—英语版本—
一开始是突触
大约 100 十亿个神经元通过“突触”相互连接,以形成人脑的各种神经回路. 虽然突触的数量可以达到 1,000 兆, 如何在电路中实际形成突触- 和神经元特异性的方式仍然不完全理解. 最近的成像研究表明,突触不仅是由遗传信息形成的, 但也通过一生的神经元活动不断修改. 此外, 越来越清楚的是许多神经发育, 神经精神病和神经系统疾病, 例如自闭症谱系障碍, 精神分裂症和阿尔茨海默氏病, 是由突触功能缺陷引起的. 所以, 必须阐明如何形成突触是至关重要的, 维护和修改以更好地了解我们的大脑在生理和病理条件下如何功能和功能障碍, 分别. 朝着这个目标, 我们关注的两个主要项目如下:
我们研究什么?
1) 神经元活动功能突触连通性的变化
突触连通性通过神经元活动的变化在功能上修改, 一种称为突触可塑性的现象. 长期增强 (LTP) 和长期抑郁症 (有限公司) 兴奋性神经传递已被深入研究,作为介导学习和记忆的突触可塑性模型. 虽然已经很明显LTP和LTD主要是由于活动引起的突触后AMPA型谷氨酸受体的增加和减少引起的 (ampars) 直接介导快速兴奋性神经传递, 分别, 尚不完全清楚地与脑功能有因果关系,与大脑功能有关,介导了体内的特定行为变化. 解决这个问题, Yuzaki实验室正在开发光遗传学工具,该工具可以直接调节活物中的LTP和LTD.
2) 突触组织者的结构突触连通性的变化
调节突触形成的分子, 维护和修改称为突触组织者. 他们分为两个主要课程, 细胞粘附分子, 例如神经蛋白和神经素, 并分泌的扩散因素, 例如Wnt和FGF. 有趣的是, 某些分泌的因素保留在突触裂缝处的细胞外基质中,并作为pre的细胞外支架- 和突触后蛋白, 因此被称为细胞外支架蛋白 (ESPS). 突触组织者的ESP型包括神经元五肽和C1Q家族蛋白, 其特征是存在C末端球状C1Q域. C1Q, C1Q家族的原型, 属于补充的先天免疫系统,并认识到各种异物. Yuzaki实验室发现C1Q家族蛋白, 例如小脑 (cblns) 和C1Q样蛋白 (C1QLS) 主要在中枢神经系统中表达,并且一直在表征其在突触时的功能.
—
最近的代表论文 (自从 2005 ):
有关详细信息:  https://orcid.org/0000-0002-5750-3544
https://orcid.org/0000-0002-5750-3544
- Kingawa W *, Paterini关闭, 松本K。, 伊莎贝尔AM, iida i, miura e, Nozawa K, Yamasaki t, Sakimura K, Yuzaki M **, 长的. 海藻酸盐受体通过与小脑的突触组织者形成复合物来调节突触完整性和可塑性. 细胞报告 43:114427, 2024.
- Itoh m, Piot l, Monly L, Paolotti P *, Yuzaki M.*缺乏GLUD受体直接配体门控离子通道活性的证据 美国科学院校 121:E2406655121, 2024.
- Nozawa K, sogabe t, Hayashi a, Motohashi J, miura e, Arai i, Yuzaki M*. 神经胶质配体的体内纳米那镜景观,依次突触规范. 神经元 110:3168-3185, 2022..
- Ojima K†, 金卡瓦W†, Yamasaki t, miura和, Itoh m, Michibata y Michibata和, Kubota r, 杜拉, miura e, Naona h, 美津, 高桥, Yuzaki M*, hamachi i *, kiyonaka s **. Coordination chemogenetics for activation of GPCR-type glutamate receptors in brain tissue. 常见的nat. 13(1):3167, 2022.
- 铃木K†, Elegheert J†, Song I†, Sasakura H†, SenkovO。, 松本K。, murning w, 克莱顿AJ, Chang VT, Ferrer-Ferrer m, miura e, Kaushik r,Ikeno M, 莫里亚, Takeuchi Y, Shimada T, 额头s, Stoyanov S., Watanabe M, Takeuchi k, dityatev a*, Aricescu ar*, Yuzaki M ** . A synthetic synaptic organizer protein restores glutamatergic neuronal circuits. 科学 369:EABB4853, 2020. (†联合第一, *共同对应, **主要作者)
- 传播k, kono m, Narumi S., Motohashi J, murning w, kohda k, Yuzaki m. Activity-dependent secretion of synaptic organizer Cbln1 from lysosomes in granule cell axons. 神经元 102:1184-1198, 2019.
- kono m, murning w, 吉田, Yuzaki m. NMDA受体中的神经元中的长期抑郁和运动学习. J生理学. 597:903-920, 2019.
- murning w, kahoh a, Narumi S., miura e, Motohashi J, 高桥a, kohda k, 福川Y, Yuzaki M **, 松本S*。 突触AMPA受体内吞作用的光遗传学控制揭示了LTD在运动学习中的作用。神经元 99:985-998, 2018. (*共同对应的作者; **主要作者).
- Yuzaki m. 中枢神经系统中的两类分泌的突触组织者. Annu Rev Physiol80:243-262, 2018.
- 萨克t, Yamashita k, Hayashi k, Miyatake s, Tamura-Nakano m, 两个h, Furuta和, Shioi g, miura e, takeo yh, 吉田, Yahikozawa h, 松本n, Yuzaki m, 铃木. MTCL1在维持Purkinje神经元轴突初始段中起着至关重要的作用. Embo j 36:1227-1242, 2017.
- Wakayama s, Kiyonaka s, Arai i, murning w, 松本, 传播k, Nemoto YL, Kusumi a, Yuzaki m, 哈马奇. 用于可视化活神经元中天然AMPA受体的化学标记. 共同的性质 8:14850, 2017.
- Yuzaki m, Aricescu ar. 一个充满成年的故事. 趋势神经科学 40:138-150, 2017.
- 额头s, Konno K., 安倍, Motohashi J, kohda k, Sakimura K, Watanabe M, Yuzaki m. CBLN1在小鼠非运动功能中的作用. J Neurosci, 36:11801-11816, 2016.
- Elegheert J, murning w, 粘土是, 小腿n, 又来了, 松本K。, kohda k, miura e, 罗斯曼, Mindakidis n, Motohashi J, Chang VT, Siebold c, Greger IH, 中川T, Yuzaki M*, Aricescu ar*. 集成GLUD受体在突触组织器中的结构基础. 科学 353:295-299, 2016. (*共同对应作者).
- Kiyonaka s, Kubota r, Michibata y Michibata和, Sakakura m, 高桥h, 命名为t, inoue r, Yuzaki m, 哈马奇. 使用活细胞内的配位化学对膜结合的谷氨酸受体的变构激活. 自然化学 8 :958-967, 2016.
- 松本K。, budentoso t, Mindakidis n, Suy, miura e, murning w, Yamasaki m, Konno K., Uchigashima m, 安倍, 渡边i, 卡诺, Watanabe M, Sakimura K, Aricescu ar, Yuzaki m. C1Q样蛋白的海藻酸盐受体功能的反式突触调节. 神经元 90:752-767, 2016.
- takeo yh, murning w, miura e, Yuzaki m. RORα调节体内小脑Purkinje细胞中树突发育的多个方面. J Neurosci 35:12518-12534, 2015.
- murning w, Mindakidis n, miura e, 安倍, 松本K。, takeo yh, kohda k, Motohashi J, 高桥a, 命中S., Muramatsu Si, Watanabe M, Sakimura K, Aricescu ar, Yuzaki m. 为了确定和维持小鼠小脑中的单打攀岩纤维,需要顺行C1QL1信号传导. 神经元 85:316-329, 2015.
- 松本, murning w, budentoso t, 野村, kohda k, Yuzaki m. Stargazin在长期抑郁症中调节通过衔接蛋白复合物调节AMPA受体运输. 共同的性质 4:2759, 2013.
- kohda k, murning w, 松本, Yamamoto t, Hirano H, Yuzaki m. D2谷氨酸受体大门长期抑郁,通过协调两个AMPA受体磷酸化位点之间的相互作用. 美国科学院校 110:E948-957, 2013.
- ito-ishida a, 宫崎T, miura e, 松本K。, Watanabe M, Yuzaki M*, Okabe S*. 突触前释放的CBLN1在小脑突触形成过程中与GLUD2相互作用会诱导动态轴突结构变化。神经元 76:549-564, 2012. (*共同对应作者).
- Unoki t, 松本, murning w, 来自UT, kohda k, 铃木A。, funakoshi y, 长谷川h, Yuzaki M*, 你好. NMDA受体介导的PIP5K激活产生PI(4,5)P2对于LTD期间的AMPA受体内吞作用至关重要。神经元 73: 135-148, 2012. (*共同对应作者)
- murning w, Miyoshi y, 哈马斯, k, 松本, 松本K。, kohda k, 我, Motohashi J, Konno r, Zaitsu k, Yuzaki M. D-丝氨酸通过Delta2谷氨酸受体调节小脑有限公司和运动配位。自然神经科学 14: 603-611, 2011.
- Yuzaki M. Cbln1及其家族蛋白在突触形成和维护中。Curr Opin Neurobiol21:215-220, 2011.
- Jitsuki s, tamemoto, 川崎T, 塔达h, 高桥a, Becamel c, 健康, Yuzaki m, Zukin RS, Ziff EB, 凯塞尔(Kessels)HW, 高桥T. 5-羟色胺介导皮质回路的跨模式重组。神经元 69: 780-92, 2011.
- 松本K。, miura e, Miyzaki t, murning w, 我, Narumi S., 福川Y, ito-ishida a, Kondo t, shigemoto r, Watanabe M, Yuzaki m. CBLN1是孤儿谷氨酸受体Δ2的配体, 双向突触组织者。科学 328: 363-368, 2010.
- Nishiyama J, 松本K。, murning w, Yamada n, Motohashi J, mizushima n, Yuzaki m. 在Lurcher小鼠中对神经退行性的重新评估: 组成型离子通量导致细胞死亡, 不是, 自噬。J Neurosci 30:2177-87, 2010.
- kakegawa, W., 宫崎骏, T., 点, K., 马苏达, K., EMI, K., Motohashi, J., 渡边, M., 浅, m. Glud2的N末端结构域 (glurΔ2) 招募了突触前末端并调节体内小脑突触发生, J Neurosci 29:5738-5748, 2009.
- iijima, T., EMI。, K., 浅, m. CBLN1表达的活动依赖性抑制: 小脑突触的发育和稳态调节机制。j. Neurosci29:5425-5434、2009.
- 松本, miura e, 松本K。, murning w, kohda k, Watanabe M, Yuzaki m. 缺乏衔接蛋白AP-4的神经元轴突自噬小体中AMPA受体的积累。神经元 57: 1-16, 2008.
- ito-ishida, 一个。, Miura, E., EMI, K., 马苏达, K., iijima, T., Kondo, T., 点, k. 渡边, M., 浅, m. CBLN1调节成熟小脑Purkinje细胞中兴奋性突触的快速形成和维持体外 和徒然. J Neurosci 28:5920-5930, 2008.
- kakegawa, W., 宫崎骏, T., EMI, K., 马苏达, K., 点, K., Motohashi, J., 机器, M., 卡瓦哈拉, s, 渡边, M., 浅, m. Glurdelta2的C末端PDZ结合基序对突触可塑性和小脑运动学习的差异调节。J Neurosci 28:1460-1468, 2008.
- kakegawa,W., 点, K., 浅, m. Delta2“离子型”谷氨酸受体充当非离子受体,可控制小脑突触可塑性。J生理学 584:89-96, 2007.
- kakegawa, W., 宫崎骏, T., hirai, H., Motohashi, J., 机器, M., 渡边, M., 浅, m. 通道孔的Ca2+通透性对于Delta2谷氨酸受体调节突触可塑性和运动配位并不是必需的。J生理学 579.3:729-735, 2007.
- hirai h, pang z, bao d, 宫崎T, 那, miura e, 帕里斯J。, y, 渡边w, Yuzaki M*, 摩根吉. CBLN1对于小脑的突触完整性和可塑性至关重要。自然神经科学 8: 1534-1541, 2005. (*共同对应作者)